

Анатомо-гистологическое строение органов пищеварительной системы диких птиц
Анализ литературных источников даёт общее представление о морфологии пищеварительной системы отдельных представителей класса Aves (Воронов, 1973, 1978, 1983; Аманова, 1978, 2002; Богатырь, 1983; Могильная, Богатырь, 1983; Richardson, Woller, 1988; Аманова, Нияздурдиева, 1989; Замосковский, 1989; Мамедова, 1991, 2002; Piersma et al., 1993; Kloss, Starck, 1994; Аманова, Мамедова, 1995; Hume, Biebaach, 1997; Бобылёв и др., 2002; Скаредова и др., 2005). Целью наших исследований было выяснить общие закономерности и особенности морфофункциональной организации пищеварительной системы представителей класса Aves с различной трофической специализацией.
Для этого было необходимо решить следующие задачи:
- исследовать в сравнительном аспекте анатомическое и гистологическое строение пищеварительной системы птиц с различными вариантами трофической специализации;
- определить защитные структуры пищеварительной системы птиц;
- проанализировать общие закономерности морфо-функциональной организации пищеварительной системы птиц в аспекте приспособлений к различным типам питания, полёта и кормодобывания.
ЗАКОНОМЕРНОСТИ МОРФО-ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ ПТИЦ с РАЗЛИЧНой ТРОФИЧЕСКой СПЕЦИАЛИЗАЦИеЙ: АНАТОМО-ГИСТОЛОГИЧЕСКОЕ СТРОЕНИЕ ОРГАНОВ ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ ДИКИХ ПТИЦ/
Л.П. Харченко, М.Ф. Ковтун Харьковский национальный педагогический университет им. Г.С. Сковороды, ул. Артёма, 29, Харьков, 61002, Украина Ин-т зоологии им. И.И. Шмальгаузена НАН Украины, ул. Богдана Хмельницкого, 15, Киев, 61601, Украина
Материал и методы исследования
Материал для исследования был собран в общих экспедиционных выездах с орнитологами Ставропольского и Ростовского государственных университетов Российской Федерации в 1990–2002 гг., а также в экспедиционных выездах в Золочевский, Чугуевский, Змиевской и Зачепиловский р-ны Харьковской обл. в 1994–2003 гг. Были использованы также фондовые материалы Музея природы Харьковского национального университета имени В.Н. Каразина и Зоологического музея Харьковского национального педагогического университета имени Г.С. Сковороды.
Проведены комплексные морфологические исследования пищеварительной системы у 340 особей 74 видов птиц разных трофических групп из 33 семейств и 15 отрядов класса Aves: поганкообразные Podіcіpedіformes; пеликанообразные Pelecaniformes; аистообразные Ciconiiformes; гусеобразные Anserіformes; соколообразные Falconіformes; курообразные Gallіformes; журавлеобразные Gruіformes; ржанкообразные Charadrіiformes; голубеобразные Columbіformes; совообразные Strіgіformes; стрижеобразные Apodіformes; козодоеобразные Caprimulgiformes; ракшеобразные Coracіiformes; дятлообразные Pіcіformes; воробьинообразные Passerіformes.
Исследование анатомического строения и морфометрических показателей пищеварительной системы птиц проводили на свежем, замороженном или фиксированном в 6–8% растворе нейтрального формалина материале. Макрорельеф внутренней поверхности стенки пищевода, желудка и кишечника исследовали на фиксированных препаратах с использованием стереоскопического микроскопа МБС-10. При фотографировании использовали цифровой фотоаппарат “Samsung”.
Гистологические исследования проводили в соответствии с общепринятыми методиками, изложенными в руководствах по гистологической технике (Лилли, 1969; Меркулов, 1969; Саркисов, Петров, 1986). Общее гистологическое строение стенки пищеварительного тракта изучали на поперечных и продольных срезах, сделанных в разных его участках. Морфометрию оболочек стенки пищеварительного тракта исследовали с использованием окуляр-микрометра АМ-9-2. В процессе изучения гистологического строения пищеварительной системы птиц разных трофических групп были изготовлены более 700 гистологических препаратов.
Гистохимические методы исследования применяли для выявления ряда биологически активных веществ и их локализации:
- секрецию полисахаридобелков в составе секретов бокаловидных клеток и клеток поверхностного эпителия железистого желудка, которые продуцируют слизь (метод окрашивания PASреакция);
- локализацию хлоридов в составе секреторных отделов сложных желёз железистого желудка (метод окрашивания конго красным);
- для выяснения происхождения секрета трубчатых желёз железистого желудка, сведения о характере и биологическом значении которого в литературе отсутствуют, и для дифференциации мышечных и соединительнотканных элементов стенки пищеварительной трубки был использован метод окрашивания фуксин-анилин-блау-оранжем по Маллори.
Материал для иллюстраций получен с использованием микроскопа “Jenamed” фирмы “Carl Zeіss” и фотоаппарата “Canon Power Shot G5”.
Результаты и обсуждение
Анатомия органов пищеварительной системы птиц разной трофической специализации
Общий план анатомической организации пищеварительной системы представителей класса Aves демонстрирует сходство с таковым представителями классов Reptilia и Mammalia, объединяя в своём строении анцестральные и продвинутые признаки. Пищеварительный тракт рептилий, птиц и млекопитающих состоит из трёх основных отделов:
- передняя кишка;
- средняя кишка;
- задняя кишка.
Основные особенности пищеварительного тракта птиц: укороченный кишечник; слабая морфологическая и функциональная дифференцировка кишечника на отделы.
Толстый кишечник у птиц представлен только прямой кишкой. Всасывание питательных веществ осуществляется практически по всей длине кишечника благодаря наличию ворсинок.
Пищевод.
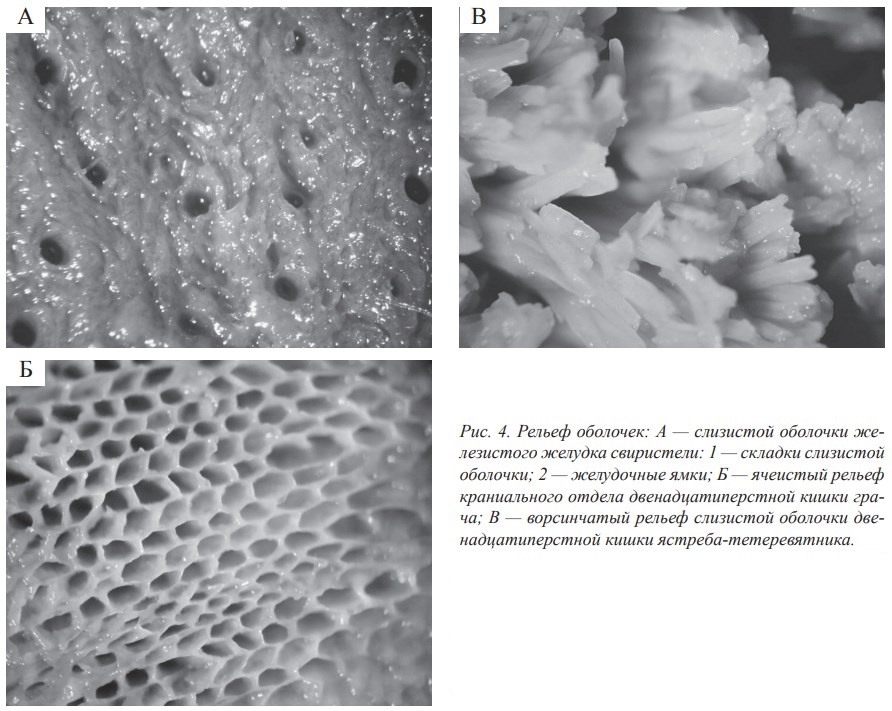
Длина пищевода зависит от длины шейного отдела позвоночника птиц и поэтому весьма изменчива в пределах класса. Складчатость слизистой пищевода присуща птицам всех трофических специализаций. При этом складки отличаются широким морфологическим и топографическим многообразием. Мы выделяем следующие типы складок: по структуре — сплошные и фрагментарные; по направленности — продольные и поперечные; по форме — ребристые, извитые, волнообразные, ветвистые, наплывающие и гофрированные; по размерам — высокие, широкие и низкие (рис. 1, А, Б).
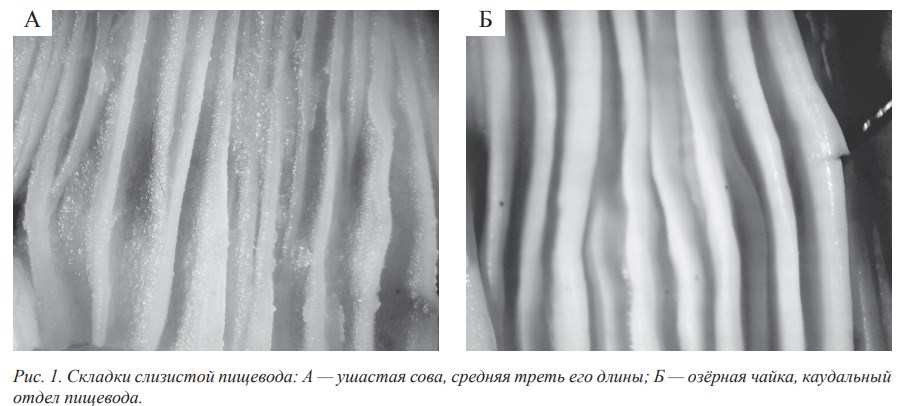
Наши данные свидетельствуют о достаточно высокой вариабельности складчатости слизистой оболочки стенки пищевода у представителей разных трофических групп. По этому признаку прослеживается также достаточно заметная внутривидовая изменчивость. Установлено, что основными факторами, обусловливающими складчатость и сложность рельефа слизистой оболочки стенки пищевода, являются наличие или отсутствие зоба и размеры пищевых объектов. Особенностью слизистой оболочки пищевода птиц является отсутствие или слабое ороговение эпителия слизистой оболочки.
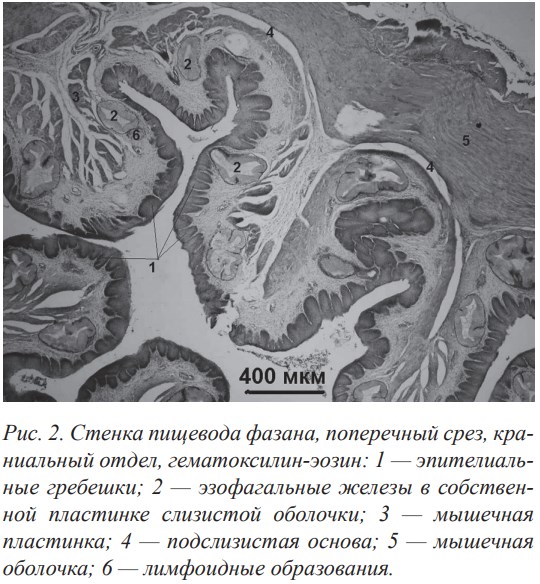
Очевидно, слабое ороговение эпителиального слоя слизистой оболочки стенки пищевода компенсируется значительным развитием железистого аппарата, который выполняет как функцию смачивания корма, так и механической защиты. Установлена зависимость между степенью развития железистого аппарата в стенке пищевода и типом питания. Питание насекомыми с твёрдым хитиновым покровом или сухим и твёрдым растительным кормом способствовало формированию хорошо развитого железистого аппарата в слизистой оболочке стенки пищевода (рис. 2). Обилие слизистых желез, лимфоидных элементов, складчатость, наличие зоба у некоторых видов птиц позволяет утверждать,что функция пищевода птиц не ограничивается только транспортировкой пищи.
Желудок.
Основными параметрами или признаками, по которым мы сравнивали строение желудка у птиц разных трофических специализаций, были: камерность; форма; строение и макрорельеф слизистой оболочки; наличие слепых мешков; толщина стенок мышечного желудка.
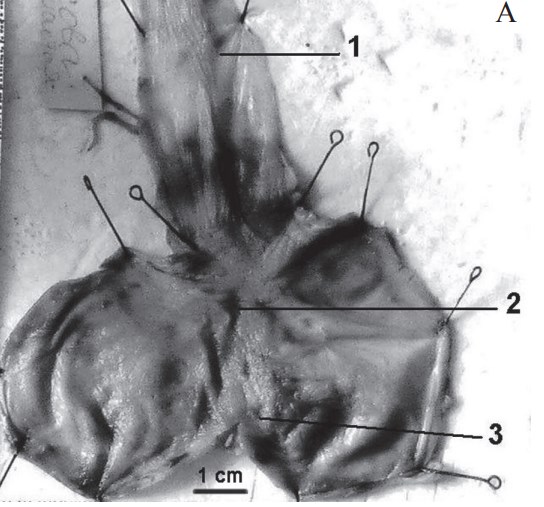
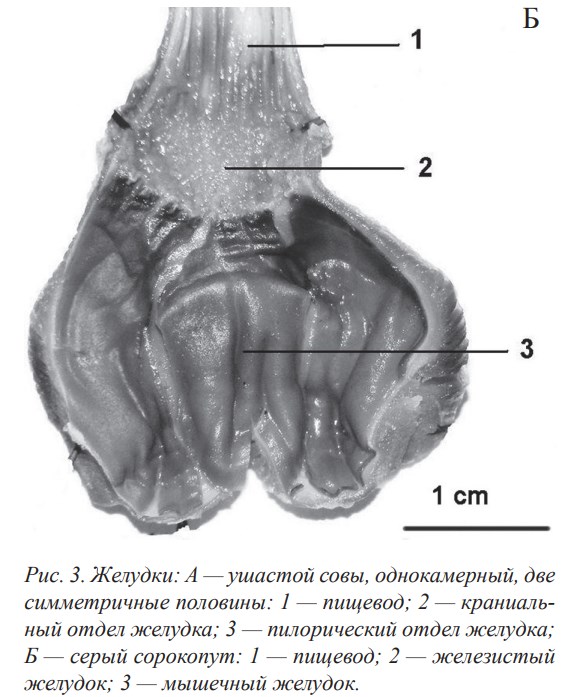
По признаку камерности желудка всех исследованных птиц можно разделить на две группы:
- Птицы, обладающие анатомически однокамерным желудком железистого типа. Такой желудок может иметь хорошо развитый пилорический мешок или не иметь его. Наличие пилорического мешка характерно для представителей отрядов аистообразных (Ciconiiformes) и пеликанообразных (Pelecaniformes). Макроморфологические исследования пилорического мешка показали, что вход и выход из него регулируются запирательными слизисто-мышечными кольцевыми складками, что способствует порционному поступлению корма и является приспособлением для более длительной её задержки в желудке (Харченко, Коц, 2001). Отсутствие пилорического мешка наблюдается у представителей отрядов соколообразных (Falconiformes) и совообразных (Strigiformes, рис. 3, А). Подчеркнём, что группа плотоядных птиц очень неоднородна по анатомическому строениию желудка.
- Для второй группы птиц характерен анатомически выраженный двухкамерный желудок (камеры называют железистым и мышечным желудками), который имеют представители всех остальных исследованных отрядов класса Aves (рис. 3, Б).
Железистый желудок имеет относительно небольшие размеры и составляет по нашим данным 0.1–0.5% от общей массы тела. Сложные глубокие трубчато-альвеолярные железы, вырабатывающие пепсиноген и соляную кислоту (у птиц, в отличие от млекопитающих, эти железы не дифференцированы на главные и обкладочные), расположены в подслизистой основе слизистой оболочки. Характерным признаком слизистой оболочки железистого желудка является складчатость и наличие отверстий выводных протоков желёз (рис. 4, А). Форма железистого желудка не отличается большим разнообразием; количество выводных протоков желудочных желёз на единицу площади у представителей различных трофических специализаций изменчиво, а их глубина зависит от типа питания.
Мышечный желудок по размерам и форме разнообразнее, чем железистый. Его масса среди исследованных видов птиц колеблется в пределах 1.5–10.0% от массы тела. Обнаружены внутривидовые вариации формы мышечного желудка, проявляющиеся чаще всего у всеядных птиц. Размеры и форма мышечного желудка зависят от трофической специализации. Особенностью строения мышечного желудка птиц является наличие слепых мешков, которые обеспечивают увеличение его объёма за счёт эластичности стенок при механической обработке корма.
Слизистая оболочка мышечного желудка покрыта кутикулой, которая имеет послойное строение. Наиболее чёткая слоистость кутикулы обнаружена нами у растительноядных и всеядных птиц, тип питания оказывает влияние и на толщину кутикулярного слоя. Так, у питающихся богатыми клетчаткой растительными кормами птиц кутикула наиболее твёрдая и толстая; среди насекомоядных такую кутикулу имеют птицы, которые питаются насекомыми с твёрдым хитиновым покровом или насекомыми с жалящим аппаратом. Мышечный желудок характеризуется значительной толщиной стенки, которая состоит из четырех мышц (две промежуточные, вентральная и дорсальная). У исследованных плотоядных птиц толщина стенки желудка заметно меньше в сравнении с растительноядными.
Кишечник.
Известно, что кишечник птиц укорочен и относительно слабо дифференцирован на отделы по сравнению с таковым у млекопитающих. Тем не менее, на фоне укороченности кишечника сохраняется общая для позвоночных закономерность: растительноядные виды имеют более длинный кишечник по сравнению с плотоядными. Толстый и тонкий отделы кишечника практически не дифференцированы. В тонком кишечнике относительно хорошо различается двенадцатиперстная кишка и почти отсутствует дифференциация на тощую и подвздошную кишки.
Полученные результаты свидетельствуют о том, что такие признаки как общая длина кишечника, относительная длина его отделов, число петель кишечника, их топография, а также рельеф слизистой оболочки определяются несколькими факторами, главным среди которых является, очевидно, тип питания (трофический фактор). К другим факторам можно отнести особенности кормодобывательного стереотипа, величину и массу птицы, особенности полёта.
По нашим данным, у растительноядных птиц общая длина кишечника превышает длину тела (от 1-го шейного позвонка до конца копчика) в 6–9 раз; у плотоядных — в 1.5–2.5 раза; у насекомоядных — в 3–3.5 раза; у зерноядных — в 3–4 раза; у всеядных — в 3–5 раз. Таким образом, у насекомоядных и плотоядных птиц, кормодобывательный стереотип которых состоит в добыче корма в полёте (ласточка-береговушка Riparia riparia, чёрный стриж Apus apus, золотистая щурка Merops apiaster), кишечник на 10–20% короче, чем у птиц-собирателей, имеющих такие же размеры и массу тела. Более того, у стрижа кишечник укорочен в большей степени, чем у щурки, что, видимо, связано с различной степенью атакующего компонента при добывании корма в полёте.
В одной и той же трофической группе птиц, но при разных размерах и массе тела, относительная длина кишечника также различна. У птиц с большими размерами и массой тела кишечник относительно длиннее. Здесь, очевидно, можно применить правило соотношения поверхности и объёма (Шварц и др., 1968), когда для поддержания необходимой эффективности обработки пищи происходит увеличение не только абсолютной, но и относительной длины кишечника.
Нами установлено, что размеры двенадцатиперстной кишки также коррелируют с трофической специализацией птиц. Наибольшая относительная длина двенадцатиперстной кишки отмечена у плотоядных и насекомоядных птиц — соответственно, 22 и 34%. Наименьшая — у растительноядных: от 5 до 9%. У зерноядных она составляет 10–17%, у всеядных — 20–25%.
Как показали наши исследования, толстый кишечник относительно удлинён у растительноядных птиц и составляет 12–16% от общей длины кишечника. У птиц с атакующей стратегией добывания корма (ласточкабереговушка, чёрный стриж) длина толстого кишечника составляет 2–5% общей длины кишечника. М.Б. Аманова (1977, 1981) выявила зависимость длины толстого кишечника от условий обитания птиц. У птиц, обитающих в аридных условиях, относительная длина толстого кишечника достигает 15%.
Интересно, что длина парных слепых кишок ушастой совы (Asio otus) составляют 17.5% от общей длины кишечника. У других представителей плотоядных птиц слепые кишки рудиментарны. Естественно, что наиболее развиты слепые кишки у растительноядных видов. По нашим даным, у белой куропатки (Lagopus lagopus) их длина достигает 52% общей длины кишечника; у обыкновенного фазана (Phasianus colchicus) — 30%.
Исследование рельефа слизистой оболочки кишечника птиц разных трофических групп позволило выделить два типа рельефа: ворсинчатый и пластинчатый. Ячеистый и лабиринтовый типы рельефа слизистой оболочки мы рассматриваем как разновидность пластинчатого. Ворсинчатый тип рельефа слизистой оболочки характерен для плотоядных птиц; пластинчатый (лабиринтовый) — для растительно- и зерноядных птиц; пластинчато-ячеистый — для всеядных птиц; ворсинчато-пластинчатый — для насекомоядных птиц (рис. 4, Б, В). Ворсинки слизистой оболочки стенки кишечника характеризуются неправильной полиморфной формой с рассеченными верхушками, что способствует увеличению поверхности пищеварения и всасывания при относительно коротком кишечнике. Усложнение пластинчатого типа рельефа (лабиринты и ячейки), очевидно, способствует задержанию химуса для пролонгации его контакта с пищеварительными ферментами, что важно при относительно коротком кишечнике. Для всех типов рельефа кишечника птиц характерна общая закономерность — в каудальном направлении уменьшается высота и увеличивается ширина ворсинок и пластинок, а глубина ячеек уменьшается.
Анализ данных анатомических исследований стенки пищеварительного тракта птиц свидетельствует об изменчивости (межвидовой и внутривидовой) отдельных признаков и о консерватизме других, что имеет определенный биологический смысл и, в частности, способствует адаптивной пластичности пищеварительной системы птиц к кормам разного происхождения и калорийности. Особенности гистологического строения органов пищеварительной системы птиц с различной трофической специализацией
Как уже было сказано, слизистая оболочка стенки пищевода птиц выстлана многослойным плоским эпителием, который способен к разной степени ороговения, что связано с кормовой специализацией. Пищеводные железы у всех исследованных птиц образованы призматическим однослойным эпителием, как и у пресмыкающихся, кроме черепах. У млекопитающих они выстланы многослойным эпителием. У птиц между секреторными отделами желёз хорошо развита капиллярная сетка сосудов. Нами установлено, что возле желёз концентрируются отдельные миоциты или их пучки, сокращение которых способствует выделению секрета (особенность, характерная для представителей класса Aves). У большинства исследуемых птиц вблизи желёз диффузно располагаются лимфоциты и лимфоидные узелки (рис. 5, А).
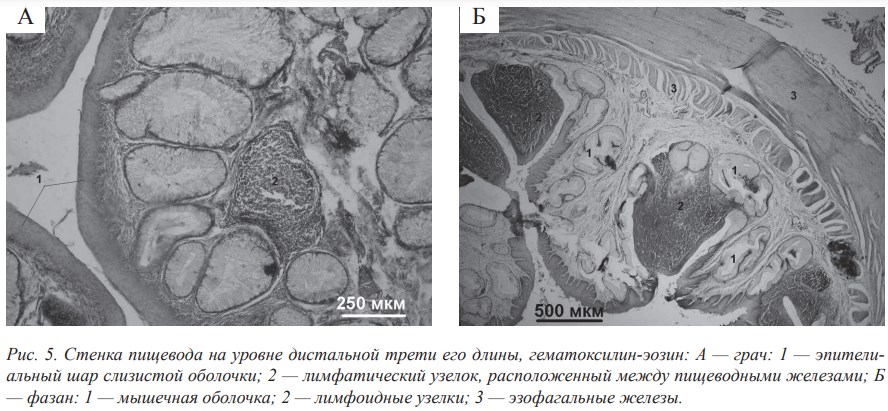
Мышечная оболочка в стенке пищевода большинства исследованных птиц хорошо развита и состоит из гладкой мышечной ткани, между миоцитами которой залегают прослойки рыхлой соединительной ткани. Взаимное расположение внешнего и внутреннего слоёв мышечной оболочки у птиц аналогично таковому у пресмыкающихся. Характерная особенность строения мышечной оболочки стенки пищевода птиц, связанна с наличием зоба, состоит в изменении направления расположения миоцитов в слоях мышечной оболочки в участках до и после зоба (рис. 5, Б).
Среди гистологических особенностей строения стенки пищевода птиц, необходимо отметить особый механизм защиты слизистой оболочки пищевода у насекомоядных птиц, которые питаются насекомыми с жалящим аппаратом (например, у золотистой щурки). Сосуды микроциркуляторного русла в слизистой оболочке стенки пищевода образуют две хорошо развитые сетки: одна — поверхностная — разветвляется непосредственно под базальной мембраной эпителия и между секреторными отделами желёз; вторая — глубокая — на границе подслизистой основы слизистой и мышечной оболочек. При этом в хорошо развитой подслизистой основе пищевода золотистой щурки сосуды микроциркулярного русла почти не определяются. Мы полагаем, что слабое развитие микроциркуляторного кровеносного русла слизистой оболочки пищевода золотистой щурки предотвращает возможность всасывания токсинов в кровь, что чрезвычайно важно в случае проникающего действия жала.
В зоне перехода пищевода в железистый желудок эпителиальный слой слизистой оболочки утончается. В собственной пластинке слизистой оболочки появляется большое количество лимфоидно-ретикулярной ткани, которая формирует в этой зоне пищеводную миндалину (рис. 5, Б).
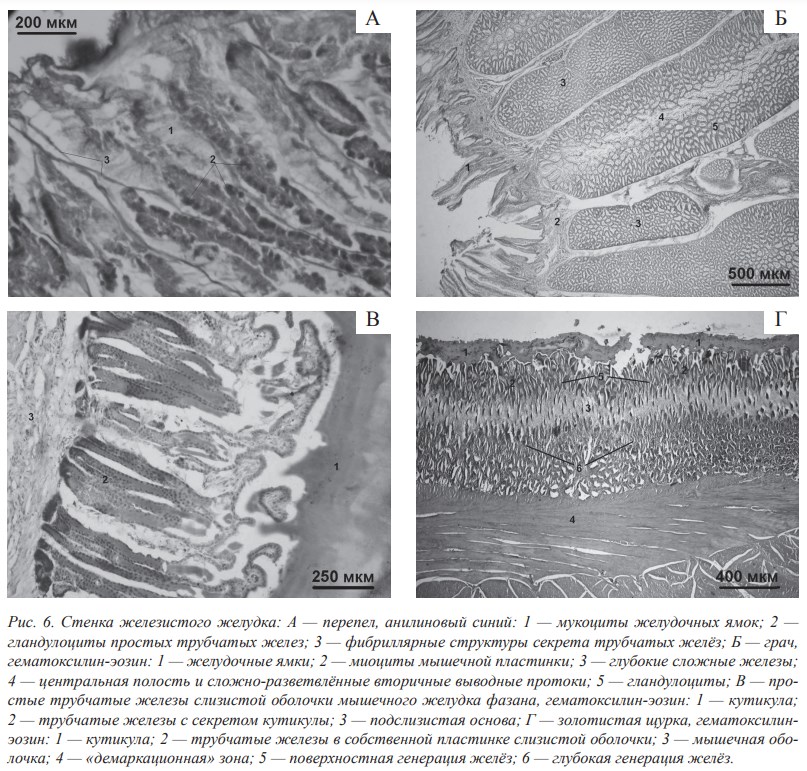
Слизистая оболочка железистого желудка у исследованных видов птиц выстлана однослойным призматическим железистым эпителием и образует щелевидные впячивания — желудочные ямки, в которые открываются протоки простых трубчастых неразветвлённых желёз. К особенностям микрорельефа слизистой оболочки стенки железистого желудка птиц необходимо отнести наличие очень глубоких желудочных ямок в собственной пластинке слизистой оболочки. В трубчатых железах стенки железистого желудка и на поверхности слизистой оболочки у всех исследованных нами птиц впервые обнаружен секрет, характерной особенностью которого является способность к образованию фибриллярных структур (рис. 6, А). Вероятно, этот секрет увеличивает плотность слизи, повышая её протективные свойства относительно механического действия корма, который попадает в железистый желудок без первичной механической обработки.
Образование желудочного сока в железистом желудке птиц обусловлено хорошо развитым секреторным аппаратом, который представлен глубокими сложными железами, расположенными в подслизистой основе слизистой оболочки. Характерной особенностью сложных желёз слизистой оболочки железистого желудка птиц является сложная система выводных протоков, которая включает первичные, вторичные и третичные протоки. При этом вся система выводных протоков выстлана мукоцитами (рис. 6, Б). Установлено, что мышечная пластинка слизистой оболочки стенки железистого желудка фрагментарна, отдельные её миоциты проникают между сложными железами, что способствует более эффективному выведению секрета из желёз.
Мышечный желудок — это полый орган, его слизистая оболочка выстлана однослойным призматическим железистым эпителием, впячивания эпителия в собственную пластинку слизистой оболочки образует простые трубчатые железы с оксифильным секретом. По нашим данным, у подавляющего большинства птиц эти железы являются простыми неразветвленными трубчатыми. Их секреторные отделы расположены плотно и параллельно друг другу, пронизывая почти всю толщину собственной пластинки слизистой оболочки. Выводные протоки открываются на поверхность слизистой оболочки (рис. 6, В). Внутренняя поверхность стенки мышечного желудка покрыта кутикулой. В её формировании принимают участие как железистые клетки поверхностного эпителия, так и секреторные клетки трубчатых желёз.
У всех исследованных птиц кутикула мышечного желудка исчерчена: секреция осуществляется дискретно, в результате чего в кутикуле заметны горизонтальные тёмные и светлые полосы. У птиц, которые питаются относительно мягким кормом (плотоядные, ихтиофаги), формируется тонкий слой кутикулы из секрета клеток поверхностного эпителия. У ушастой совы кутикула инкрустирована волосками шерсти, которые укрепляют её и придают ей прочность. У зерноядных птиц кутикула по структуре представлена преимущественно эозинофильними вертикальными столбиками клеток, что создает высокую степень шероховатости поверхности кутикулы и способствует эффективному перетиранию оболочек зерна. У насекомоядных птиц, особенно у питающихся насекомыми с жалящим аппаратом, была обнаружена своеобразная «демаркационная» зона, которая образована соединительной тканью собственной пластинки слизистой оболочки и разграничивает две генерации желёз — поверхностную и глубокую (рис. 6, Г). Мы считаем, что поверхностный слой собственной пластинки слизистой оболочки содержит железы, которые подлежат отторжению вместе с кутикулой. В глубоком слое собственной пластинки секреторные клетки желёз находятся в стадии активной пролиферации.
Мышечная оболочка стенки мышечного желудка у всех исследованных птиц хорошо развита и образована гладкой мышечной тканью. В средней части стенки мышечного желудка мышечная оболочка двухслойная: внутренний слой — кольцевой, внешний представлен пучками, которые имеют прямоугольную и треугольную форму с косым расположением миоцитов.
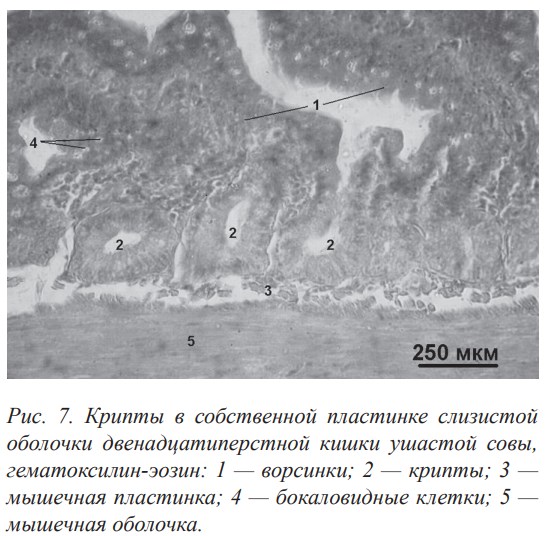
Активная поверхность слизистой оболочки стенки кишечника у птиц увеличивается за счёт сложной архитектоники рельефа слизистой оболочки и общекишечных желёз — крипт. Крипты образовались путем инвагинации поверхностного эпителия в собственную пластинку слизистой оболочки (рис. 7). В криптах кишечника птиц разных трофических специализаций обнаружены энтероциты, которые содержат положительно окрашенные гранулы (метод окраски амидочёрным В), что указывает на их белковую природу. О наличии секреторной функции клеток крипт свидетельствует также альвеолярное расширение донной части крипт, где сконцентрированы секреторные гранулы. В эпителии крипт наблюдаются отдельные окаймленные энтероциты, значительное количество бокаловидных клеток и фигуры митоза в камбиальных клетках (источник регенерации эпителия). Крипты в стенке кишечника птиц отличаются большой изменчивостью — по размерам, количеству и длине, плотности расположения, что обусловлено типом питания. Наибольшая толщина криптального слоя отмечена у плотоядных и насекомоядных птиц в краниальном отделе стенки кишечника. У птиц разных трофических специализаций в стенке кишечника между криптами были обнаружены миоциты. Наличие отмеченных выше структур (крипт) по всей длине стенки кишечника свидетельствует о том, что процессы пищеварения и всасывания осуществляются вдоль всего кишечника, и это можно рассматривать как компенсацию укороченности последнего.
Мышечная пластинка слизистой оболочки тонкого кишечника образована продольно ориентированными миоцитами, её развитие коррелирует с типом питания птиц. У всех исследованных птиц подслизистая основа слизистой оболочки слабо выражена по всей длине кишечника.
Мышечная оболочка по всей длине стенки кишечника образована двумя слоями: внешним — продольным и внутренним — кольцевым. У всех исследованных видов птиц кольцевой слой по толщине значительно превышает продольный. Соотношение толщины продольного и кольцевого слоёв составляет: у плотоядных — 1:7.5; у насекомоядных — 1:3.5; у всеядных птиц — 1:10.0; у растительноядных — 1:4.0; у зерноядных — 1:7.0. Мы считаем, что именно отмеченное выше соотношение толщины слоёв мышечной оболочки обусловливает активные перистальтические и антиперистальтические сокращения стенки кишечника.
По сравнению с тонким кишечником в толстом кишечнике, который представлен прямой кишкой, подслизистая основа слизистой оболочки разделяет мышечную пластинку слизистой оболочки и мышечную оболочку. Толщина мышечной оболочки стенки прямой кишки у всех исследованных птиц заметно увеличивается преимущественно за счёт кольцевого и в меньшей степени продольного слоёв. При этом у большинства исследованных птиц, кроме плотоядных и насекомоядных, между миоцитами мышечной оболочки обнаружены значительные прослойки рыхлой соединительной ткани, что является свидетельством способности прямой кишки к увеличению её объема, т.е. к растягиванию.
Особенности гистологического строения стенок слепой кишки птиц разных трофических групп в значительной степени предопределены наличием в собственной пластинке слизистой оболочки скоплений лимфоидной ткани, что указывает на функциональную причастность её к системе иммунной защиты пищеварительного тракта. Слепые кишки у зерноядных и всеядных птиц, а также ихтиофагов, представлены лимфо-эпителиальными комплексами. Особый интерес представляют крупные слепые кишки ушастой совы, которые составляют до 17% от общей длины кишечника. Гистологическое строение стенки слепой кишки этих птиц (наличие большого количества плотно расположенных ворсинок) позволяет утверждать, что в данном отделе кишечника интенсивно происходят процессы всасывания.
Лимфоидные образования в структурах пищеварительного тракта птиц разных трофических групп
В пищеварительный тракт птиц с кормом постоянно попадают антигены разного происхождения. Результаты наших исследований показали определённую взаимосвязь между особенностями трофической специализации птиц и степенью развития лимфоидных элементов в стенке пищеварительного тракта. У исследованных птиц обнаружена общая закономерность по локализации лимфоидных образований в стенке пищеварительного тракта — наибольшее их количество сконцентрировано в местах перехода из одного отдела пищеварительного тракта в другой. Особо большое количество лимфоидных образований сконцентрировано на границах между пищеводом и железистым желудком, а также между тонким кишечником и прямой кишкой.
Отсутствие глоточных миндалин у птиц (корм не задерживается в ротовой полости) компенсируется развитием лимфоидных структур в стенке пищевода, включая и пищеводную миндалину в участке перехода из пищевода в железистый желудок. Лимфоидная миндалина существенно отличается по относительным размерам у насекомоядных птиц с разными стратегиями охоты: у ловящих насекомых в полёте и у собирающих насекомых на почве и листьях. Большие относительные размеры лимфоидная миндалина имеет у растительноядных и зерноядных птиц, корм которых постоянно инфицирован антигенами (рис. 5, Б). У птиц, которые питаются насекомыми с жалящим аппаратом, лимфоидные структуры локализованы преимущественно в подслизистой основе слизистой оболочки и представленны в основном переваскулярными скоплениями лимфоцитов.
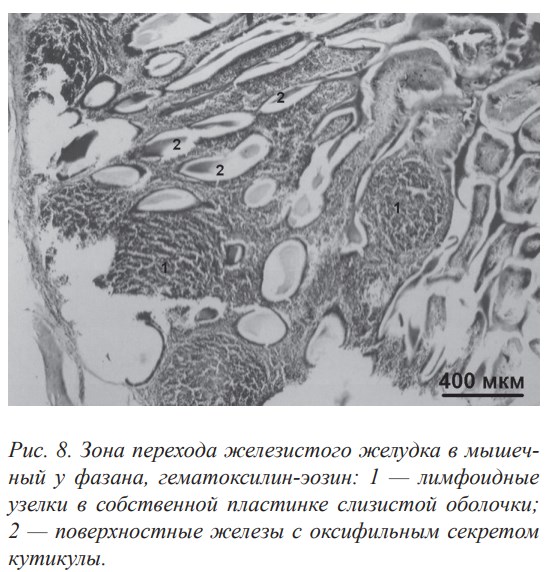
У исследованных птиц стенки железистого и мышечного желудков умеренно инфильтрованы лимфоцитами. В собственной пластинке слизистой оболочки стенки железистого желудка встречаются одиночные лимфоидные узелки овальной или округлой формы, расположенные вокруг выводных протоков сложных желёз. Большие лимфоидные образования в собственной пластинке слизистой оболочки представлены нодулярными скоплениями лимфоидной ткани и сконцентрированы в зоне перехода железистого желудка в мускульный (рис. 8). Морфофункциональная связь секреторных отделов желёз с подэпителиальными лимфоидными узелками обеспечивает образование лимфоэпителиальных комплексов. Функционально такие комплексы образуют защитный барьер за счёт способности слизи выносить и удерживать на поверхности слизистых оболочек секреторные иммуноглобулины.
У всех исследованных птиц стенка тощей и подвздошной кишок умеренно инфильтрована диффузно расположенными лимфоцитами и отдельными лимфоидными узелками. Особенностью стенки тонкого кишечника птиц является то, что все её оболочки инфильтрованы разными лимфоидными структурами.
По топографии и концентрации можно выделить следующие лимфоидные структуры:
- лимфоидная миндалина — в зоне перехода пищевода в железистый желудок;
- лимфоэпителиальные комплексы;
- лимфоидные узелки;
- лимфоидные бляшки
- узелки и диффузно расположенные лимфоциты в слизистой, мышечной и серозной оболочках;
- лимфоциты, расположенные между эпителиоцитами.
Описанные лимфоидные структуры, очевидно, составляют собственный защитный барьер пищеварительной системы, способствуя обезвреживанию токсинов и микроорганизмов, поступающих в пищеварительный тракт с необработанной пищей.
Заключение
На фоне разнообразия трофических специализаций у представителей класса Aves морфологическая организация их пищеварительной системы представляется универсальной. Однако такое впечатление складывается, когда мы рассматриваем только анатомию или гистологию (точно так же — физиологию или биохимию без учёта морфологических исследований); когда исследование проведено на отдельных видах, небольшой группе видов или на отдельных органах пищеварительной системы. Только при исследовании большого числа видов с различными трофическими специализациями начинают проявляться так называемые «частные» различия в строении тех или иных структур, в совокупности составляющих пищеварительную систему. В представленном сообщении такими «частными различиями» были: относительная длина кишечника и его отделов; камерность желудка и наличие дополнительных образований в нём; особенности эпителиальной выстилки кишечника; рельеф слизистой; структура стенки различных отделов кишечника и развитие мышечных компонентов. Следует добавить, что такие «частные различия» выявлены и при биохимических исследованиях активности пищеварительных ферментов, и при гистохимическом исследовании железистых клеток и природы их секрета.
Благодарности
Авторы приносят большую благодарность за оказанную помощь в сборе материала для исследований доктору биологических наук, профессору Ставропольского государственного университета А.Н. Хохлову; доктору биологических наук, профессору Ростовского государственного университета В.П. Белику, доктору биологических наук, профессору РГАУ и МСХА им. К.А. Тимирязева Л.В. Маловичко.
Литература
- Аманова М.Б. 1977. Некоторые морфологические особенности пищеварительного тракта представителей ткачиковых в условиях пустыни. — Экология, 1: 99–101.
- Аманова М.Б. 1978. Сравнительная морфология рельефа кишечника у представителей отряда Passeriformes. — Зоол. журн., 57 (8): 1218–234.
- Аманова М.Б. 1981. Строение стенок кишечника птиц аридных условий существования. — Орнитология, 16: 45–56.
- Аманова М.Б. 2002. Функциональная активность эпителиоцитов дистальных отделов кишечника птиц в аридных условиях существования (на примере рода Passer). — Вопросы орнитологии Туркменистана. М., 123 с.
- Аманова М.Б., Мамедова Н.М. 1995. Морфологическая адаптация пищеварительного тракта птиц различной пищевой специализации. — Экология и охрана окружающей среды. Мат-лы международной научно-практ. конф., Пермь, с. 8–9.
- Аманова М.Б., Нияздурдинова Д.К. 1989. Структурные и функциональные адаптации пищеварительного тракта к типу питания у птиц. — 7 Всесоюзная конф. по экологии и физиологии. Тезисы докладов, Ашхабад, с. 25–26.
- Бобылев А., Глотов А., Батоев Ц. 2002. Возможности пищеварительной системы птиц. — Птицеводство, 3: 14–19.
- Богатырь Л.Я. 1983. Гистохимия биополимеров функциональных структур желудка птиц. — Автореф. дис. … канд. биол. наук. М., 17 с.
- Воронов Н.П. 1973. Адаптивные особенности пищеварительной системы насекомоядных и зерноядных птиц. — Вестник зоологии, 5: 21–27.
- Воронов Н.П. 1978. Морфометрическое исследование пищеварительной системы свиристеля (Bombycilla garrulus). — Вестник зоологии, 5: 28–31.
- Воронов Н.П. 1983. Морфометрическая характеристика органов пищеварительной системы большой синицы. — Вестник зоологии, 5: 21–28.
- Замосковский Е.М. 1989. О соотношении длины отделов тонкого кишечника у птиц разного типа питания. — Межвузовский сборник научных трудов. Л., с. 167–173.
- Лилли Р. 1969. Патологическая техника и практическая гистохимия. — Пер. с англ. под ред. В.В. Португалова, М., 645 с.
- Мамедова Н.М. 1991. Сравнительно-морфологические исследования пищеварительного тракта ряда видов врановых и сорокопутовых. — Мат-лы 10 Всесоюзной орнитол. конф. Минск, с. 208–216.
- Мамедова Н.М. 2002.
Сопровождения предприятий свиноводства и птицеводства
Мы используем ветеринарные препараты нового поколения
На вашу птицефабрику или свиноводческий комплекс
Все товары и услуги сертифицированы